(Ciencias de Joseleg)(Biología)(Reproducción en los seres vivos)(Reproducción humana)(Introducción)(Historia de vida humana)(Introducción al sistema reproductor)(Regulación hormonal masculina)(Fisiología del sistema reproductor masculino)(Testosterona, masculinidad y reproducción)(La espermatogénesis humana)(Regulación hormonal femenina)(Anatomía del sistema reproductor femenino)(Los estrógenos, la feminidad y la reproducción)(La ovogénesis)(La ovulación y el cuerpo lúteo)(El ciclo menstrual)(El coito efectivo y el viaje de los espermatozoides)(De la fecundación a la implantación)(Gastrulación y formación de los discos embrionarios)(Los sacos embrionarios)(Destinos del disco trilaminar)(Gemelos y las membranas fetales)(Desarrollo fetal y embarazo)(Referencias bibliográficas)(Versión documento word)
A pesar de que los
ovarios y los testículos son órganos homólogos, su estructura interna es muy
diferente. En los testículos la gametogénesis y esteroidogénesis están
controladas por dos tipos de células separadas por la membrana de los túbulos
seminales. Sin embargo, la estructura interna del óvulo no posee dichos
túbulos, lo que tiene es una enorme cantidad de huevecillos o folículos que
contienen a los óvulos en desarrollo. A pesar de lo anterior si es posible
encontrar las dos líneas de células homólogas a las células de Sertoli y las
células de Leydig separadas por una membrana. El folículo de Graaf posee las
células teca externas que se encuentran por encima de la lámina basal, y debajo
de estas se encuentran las células granulosas. Las granulosas son homólogas a
las células de Sertoli ya que están adyacentes al óvulo, mientras que las
células teca externas son homólogas a las células de Leydig pues están
separadas por una membrana externa. Al igual que las células de Leydig, la
función de las células teca es producir testosterona y androstenodiona,
mientras que las células granulosas homólogas a las de Sertoli convierten
dichas hormonas en estradiol y estrona.
Tanto las células
granulosas y las células teca se encuentran involucradas en la síntesis y
secreción de hormonas esteroides en la mujer. El principal esteroide activo
producido por el folículo es el estradiol, un esteroide de 18 carbonos con la
misma base de la testosterona. La esteroidogénesis es el proceso de producción
de hormonas esteroides, la cual depende de la disponibilidad de colesterol, el
cual se origina por diversas rutas metabólicas, sin el colesterol como
precursor primario no es posible sintetizar las hormonas sexuales. El
colesterol ovárico se obtiene a partir de lipoproteínas solubles en la sangre,
síntesis de novo al interior de las células ováricas y esteres colesteroidales
en las gotas aceitosas al interior de las células ováricas.
Sin embargo, la
fuente la importante de colesterol para la esteroidogénesis es la lipoproteína
de baja densidad o LDL también conocido como colesterol malo por su tendencia a
acumularse en las arterias antes de ser depositado en los tejidos de destino.
La conversión de
colesterol a pregnenolona por medio del corte de la cadena lateral mediante una
enzima específica es una reacción limitante que se encuentra regulada
hormonalmente mediante la hormona luteinizante. Al igual que en las células de
Leydig, la hormona luteinizante activa una proteína G transmembranal
"figura principal" que activa un mensajero secundario intracelular llamado
AMP cíclico. Toda esta ruta aprovecha la naturaleza funcional del AMP cíclico y
los lípidos. La función basal es que el incremento de AMP cíclico es una señal
de baja energía para la célula, por lo que una respuesta lógica es que esta
incremente la absorción de lípidos, los cuales pueden ser catabolizados para
obtener energía.
En la esteroidogénesis el proceso es el mismo, solo que la
ruta metabólica en la que se involucra el colesterol no es energética, sino que
estará involucrada en la producción de hormonas. Muchas rutas metabólicas de
los seres vivos emplean secciones de otras, esto provoca un ahorro general en
la cantidad de genes involucrados en muchos procesos. La pregnenolona al igual
que en las células de Leydig se sintetiza en la mitocondria a partir del
colesterol, una vez que se ha producido la pregnenolona, esta se difunde por
transporte pasivo fuera de la mitocondria y es absorbida por el retículo
endoplasmático liso "smooth endoplasmatic reticulum", donde ocurre el
siguiente paso de la esteroidogénesis femenina.
Otro ejemplo del reciclado de rutas metabólicas se muestra en la esteroide génesis femenina. Puesto que tanto los hombres como las mujeres sintetizan testosterona y estradiol la única diferencia es la regulación para la producción diferencial de testosterona y estradiol. Las rutas intermedias para la producción de estradiol involucra la misma de la testosterona, la ruta 5 delta y la 4 delta. La pregnenolona es convertida a progesterona en la dura d4 por medio de la enzima 3b-deshidrogenasa de hidroxiesteroides, o en la ruta d5 por medio de la de la enzima 17a-hidroxilasa. En ambas rutas el producto es 17a-hidroxipregnenolona.
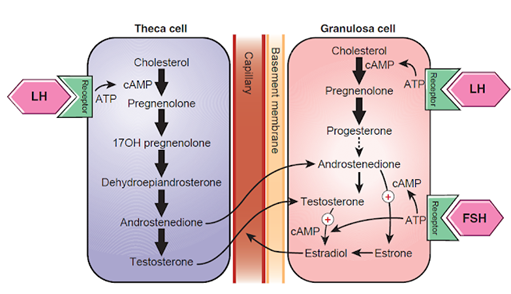
Figura 43. Comunicación hormonal paracrina entre las células teca y las
células granulosas.
La 17a-hidroxipregnenolona es convertida a androstenodiona y
testosterona por medio de la 17,20-liasa y la 17b-deshidrogenasa de
hidroxiesteroides respectivamente en la ruta d4. En la ruta d5 la
17a-hidroxipregnenolona es convertida a dehidroepiandrotestosterona por medio
de la 17,20-liasa, la cual es subsecuentemente transformada a androstenodiona
por 3b-dehidrogenasa de hidroxiesteroides. La testosterona producida se difunde
a través de las células teca, cruzan la membrana basal e ingresan a las células
granulosas.
Como se mencionó anteriormente, la ruta bioquímica para la
producción de hormonas esteroides es la misma en hombres y mujeres, sin
embargo, es la regulación fina de las hormonas la que diferencia en una mayor
producción de estrógenos que de andrógenos. El paso de la estosterona y la
androstenodiona a estradiol y estrona es realizada por la misma enzima que en
el hombre, la aromatasa. Los estrógenos poseen generalmente 18 carbonos. La
estrona puede convertirse a estradiol por la 17b-deshidrogenasa de
hidroxiesteroides.
La secreción de estradiol por parte del folículo requiere de
la cooperación de las células teca y de las células granulosas con la
coordinación de la hormona luteinizante y la hormona folículoestimulante.
Solo las células granulosas son capaces de ser activadas por
la hormona folículo estimulante, mientras que la acción de la hormona
luteinizante se extiende a las teca y las células del estroma a parte de las
mismas células granulosas. La expression de los receptors para la hormona
luteinizante es dependiente de la etapa de desarrollo del folículo, las células
teca adquieren dichos receptores temprano en su desarrollo, mientras que las
células granulosas solo expresan los dos receptores tarde en su desarrollo.
Las enzimas para las rutas bioquímicas de expresan de forma
diferencial, esto quiere decir que una sola célula es incapaz de realizar toda
la esteroidogénesis desde el colesterol hasta el estradiol. La aromatasa que
convierte los andrógenos a estrógenos solo es producida por las células
granulosas, pero estas carecen de la 17a-hidroxilasa. Esto quiere decir que las
granulosas no pueden producir los andrógenos aromatizables como la testosterona
y la androstenodiona. Son las células teca las que se encargan de producir los
andrógenos masculinizantes, los cuales se difunden hasta las células granulosas
donde son convertidos a los estrógenos feminizantes.
Al igual que las células de Leydig, las células teca se
encuentran fuera del ambiente donde se desarrollan los gametos gracias a una
membrana. Estas células se encuentran en contacto directo con capilares
sanguíneos que les suministran colesterol por medio de receptores de
lipoproteínas de baja densidad o LDL. Las células granulosas también pueden
producir testosterona, pero no por las rutas estándar de la esteroidogénesis, y
es muy poco eficiente en su producción, por lo que siempre son dependientes de
los suministros de las células teca.
La mayor población de células en el folículo son las células
granulosas, por lo que cabe de esperar que el efecto estimulante de la hormona
folículoestimulante se de en estas células, pues de allí viene su nombre. La hormona
folículoestimulante activa a las células granulosas mediante un mecanismo
dependiente de una proteína G y de AMP cíclico para aumentar la proliferación y
diferenciación de las células granulosas, inducir la producción de la enzima
aromatasa para producir los estrógenos, producir progesterona e incrementar la
producción de inhibina.
A medida que el folículo va madurando, la cantidad de
receptores para las dos gonadotropinas va aumentando. La hormona
folículoestimulante también induce que las nuevas células granulosas contengan
más receptores para sí misma y para la hormona luteinizante, haciendo que el
folículo sea cada vez más sensible a los estímulos hormonales.
La acción combinada de ambas gonadotropinas amplifica la
producción de estrógenos. Las células teca y del estroma producen hormonas
androgénicas que sirven como la base para la producción de los estrógenos. Esto
significa que, ¿si una mujer consume hormonas esteroides aromatizables como la
testosterona podra reforzar sus características femeninas al convertirlo en
estradiol? La respuesta es no.
En altas concentraciones los andrógenos son convertidos por
medio de la 5a-reductasa en la hormona esteroide androgénica más potente, la
dihidrotestosterona. La dihidrotestosterona
paraliza el desarrollo del folículo conllevando a la atresia folicular o muerte
del folículo. Si los folículos mueren rápidamente entonces la mujer sufre un
efecto masculinizante hormonal, como les ha sucedido a muchas mujeres
deportistas que han abusado de los esteroides anabólicos.
No hay comentarios:
Publicar un comentario