(Ciencias de Joseleg)(Biología)(Reproducción en los seres vivos)(Reproducción humana)(Introducción)(Historia de vida humana)(Introducción al sistema reproductor)(Regulación hormonal masculina)(Fisiología del sistema reproductor masculino)(Testosterona, masculinidad y reproducción)(La espermatogénesis humana)(Regulación hormonal femenina)(Anatomía del sistema reproductor femenino)(Los estrógenos, la feminidad y la reproducción)(La ovogénesis)(La ovulación y el cuerpo lúteo)(El ciclo menstrual)(El coito efectivo y el viaje de los espermatozoides)(De la fecundación a la implantación)(Gastrulación y formación de los discos embrionarios)(Los sacos embrionarios)(Destinos del disco trilaminar)(Gemelos y las membranas fetales)(Desarrollo fetal y embarazo)(Referencias bibliográficas)(Versión documento word)
El objetivo de la ovogénesis es la producción de óvulos capaces de ser fertilizados y convertirse en la base para un nuevo cuerpo diploide 2n mediante la combinación del contenido genético de dos células haploides n. Al igual que en la espermatogénesis, o, mejor dicho, de manera mucho más marcada los esquemas para la ovogénesis omiten una serie de detalles relacionados con los tiempos de especialización y los lugares en los que se da a cabo.

Figura 44. Ovogénesis y
foliculogénesis. La ovogénesis es más
compleja que la espermatogénesis debido a que involucra a una estructura
accesoria llamada folículo. El óvulo se desarrolla dentro del folículo, y aun
después de que ambos se separan en la ovulación, seguirán vinculados
hormonalmente. Otro detalle es que la ovogénesis solo se completa cuando el
espermatozoide lo penetra y se termina la meiosis II.
A diferencia de los espermatozoides que pasan de
espermatogonio a espermatozoide en cohortes cada 65 días, una mujer especializa
desde el ovogonio a los oocitos primarios una sola vez, durante su vida uterina
antes de nacer. Posteriormente los oocitos primarios quedarán detenidos en la
profase de la meiosis I. Posteriormente con el inicio del ciclo menstrual, cada
mes aproximadamente, los ovarios liberan un solo oocito primario que continuará
la meiosis. Adicionalmente al oocito en proceso de meiosis existe una
estructura asociada y funcional denominada folículo en el cual el oocito
primario se desarrolla. La meiosis no se termina en los ovarios, y solo hasta
que un espermatozoide ingresa en el oocito maduro “o secundario” este inicia su
meiosis II con el núcleo del espermatozoide dentro.
Los ovogonios son las células madre a partir de las cuales
se forman los óvulos maduros. Como se ha mencionado anteriormente, los
ovogonios no se regeneran como los espermatogónios, y solo se generan durante
la etapa embrionaria. Los ovogonios se generan en el saco embrionario y migran
a la cresta genital, donde se precipitan en el ovario en formación. Sin los
ovogonios los ovarios son incapaces de desarrollarse de manera correcta. Las
células germinales se dividen de manera activa por mitosis solo hasta antes del
nacimiento, cuando ingresan en el inicio de la meiosis. Los ovogonios que
inician la meiosis se denominan ovocitos primarios.
Cerca de 1 millón de ovocitos primarios ingresan en la
meiosis, pero se quedan paralizados en la profase de la meiosis I hasta la
pubertad. No todos los ovocitos primarios sobreviven la etapa de dormancia, del
millón de ovocitos primarios solo sobreviven uno 200 000 gracias a un proceso
llamado atresia. Cuando la mujer alcanza los 30 años solo sobreviven unos 26
000 y para la etapa de menopausia los ovarios están esencialmente vacíos de
ovocitos. Cada ovocito en desarrollo se encuentra rodeado por un folículo
primordial que permite el desarrollo del ovocito.
Los óvulos "izquierda Figura 44" se desarrollan al interior de una bolsa células llamada folículo "derecha Figura 44". Antes de nacer los ovogonios "oogonium" ingresan en la mitosis para diferenciarse en ovogónios secundarios. Inmediatamente después ingresan en meiosis I donde se quedan paralizados. Paralelamente los folículos se desarrollan levemente alrededor. Después de la pubertad los folículos se desarrollan alrededor de la cohorte de ovocitos liberados cada mes. Cuando finaliza meiosis I se genera una célula con casi todo el citoplasma y otra degenerada llamada cuerpo polar.

Figura 45. Modelo simplificado de la
ovogénesis/foliculogénesis. Lo que se libera del
ovario es un ovocito secundario que aún tiene que completar la meiosis, dado
que está arrestado, solo reiniciará la meiosis si un espermatozoide lo fecunda.
Todos los ovocitos sanos que se encuentran en un estado de
parálisis justo al inicio de la meiosis I en la profase. Cuando el folículo es
estimulado por hormonas de control como las gonadotropinas, el ovocito en su
interior inicia el desarrollo prosiguiendo la meiosis. Como en cualquier
proceso de meiosis, deben darse dos divisiones. Hay que recordar que los
ovocitos primarios que ingresan en la meiosis poseen un contenido genético
equivalente a 2(2n)
Cuando se da la anafase I los cromosomas homólogos son
separados, lo cual disminuye a la mitad la cantidad de cromosomas, pero cada
uno contiene dos cromátides 2(2n)
La segunda meiosis avanza hasta la metafase, donde las
células quedan paralizadas nuevamente. En este punto el óvulo maduro “oocito
secundario” es expulsado definitivamente del folículo. La segunda división meiótica se da cuando el espermatozoide penetra en
el óvulo, formando el núcleo haploide (n) que no puede
experimentar la cariogamia hasta que el núcleo del óvulo complete su última
fase miótica, por lo que tenemos un núcleo (n+n) o
dicarionte. El cuerpo polar también hace meiosis, y estos cuerpos polares
secundarios pueden generar una seria de células con función desconocida “en
otras veces el cuerpo polar se paraliza”. Cuando el núcleo del óvulo termina su
meiosis II tampoco se fusiona con el núcleo del espermatozoide, lo que va a
pasar es que ingresa a una mitosis 2(n+n) y una vez que los núcleos
de ambos gametos se dispersan, si se da la recombinación cromosómica, formando
los primeros dos blastómeros que verdaderamente son diploides (2n). Si la fertilización no se da, el ovocito
secundario comienza a degenerarse.
Cuando estudiamos la espermatogénesis y la ovogénesis
tenemos la mala tendencia de desconectar lo que sucede de los órganos que lo
permiten, lo que nos sucede para la producción de espermatozoides también es
cierto para la producción de óvulos. La producción de óvulos es marcadamente
diferente a la de espermatozoides en cuanto a la fisiología del órgano
involucrado. El óvulo no posee estructuras tubulares en donde se críen los
óvulos como sucede con los espermatozoides. De hecho, lo que tiene el ovario es
una serie de huevecillos inmaduros llamados folículos, los folículos poseen en
su interior una población celular que incluye no solo al óvulo inmaduro,
también otras células involucradas con la producción hormonal.
Los folículos son la zona que permite el desarrollo de los ovocitos primarios, y la foliculogénesis es el proceso de desarrollo y maduración de los folículos. Se asemejan a huevos que contienen una serie de células que incluyen al óvulo. Durante su desarrollo los folículos pueden presentarse en alguno de los siguientes estados: descansando y listos para ovular. Durante cada ciclo menstrual, los ovarios producen una pequeña cohorte de folículos en crecimiento y cada uno posee un óvulo en desarrollo.
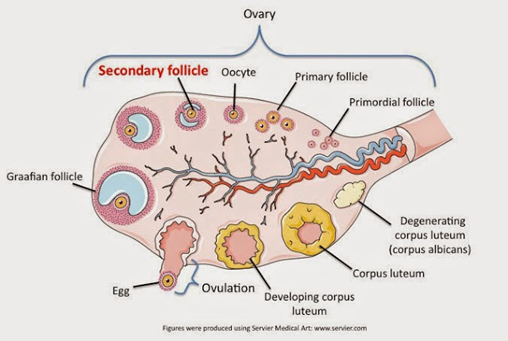
Figura 46. Modelo del ciclo folicular. Esta estructura es importante ya que segrega las hormonas femeninas, su nacimiento y muerte es lo que dicta el ciclo menstrual.

Figura 47. El folículo. Corte histológico
mostrando un folículo en formación, son estructuras extremadamente simétricas
semejantes a un embrión. lo que vemos aquí es un folículo terciario.
Maduración de un óvulo involucra la muerte de otros
Del grupo de folículos en desarrollo, solo el más fuerte
completa su proceso de crecimiento hasta liberar el óvulo, los demás son
eliminados siendo inducidos a la atresia folicular “muerte del sistema folículo
ovocito”. De toda la cohorte solo uno folículo logra desarrollarse emergiendo
como el folículo dominante, el cual ovula liberando un ovulo maduro el cual aún
no ha pasado por todas las etapas de la meiosis ovular.
Durante las etapas iniciales de la foliculogénesis, se
activas varios folículos primordiales, pero a medida que se avanza hacia los
folículos primarios la cantidad desciende, cuando se llega al folículo
secundario en cada menstruación normal solo sobrevive uno, haciendo de esto un
ejemplo de evolución darwiniana por competencia intraespecífica, solo el
folículo más fuerte logra su desarrollo.
Folículos primordiales
Los folículos primordiales son aquellas que se encuentran como reserve de óvulos a lo largo de la vida de la hembra y generalmente se considera que se encuentran en un estado de no crecimiento. La cantidad de folículos se pierde paulatinamente a lo largo de la vida del individuo femenino y para la menopausia, los ovarios están esencialmente secos de folículos. Los folículos primordiales se encuentran localizados en la corteza de los ovarios en su región periférica.

Figura 48. Corte histológico mostrando los folículos
primordiales.
Los folículos primordiales son la reserva ovárica de la
mujer, en su interior se encuentra todos los óvulos, uno por folículo. En cada
ciclo menstrual un grupo de estos es activado de forma independiente a las
gonadotrofinas, pero su desarrollo ulterior es dependiente de ellas.
La formación del folículo primario
La progresión desde los folículos primordiales a la siguiente etapa del desarrollo folicular “folículo primario” se da de manera relativamente constante desde el feto, la etapa juvenil, la prepuber y la adulta. Lo anterior implica que la formación del folículo primario es independiente de las gonadotropinas neuronales, sin embargo, sin estas hormonas es imposible que un folículo pueda madurar a las etapas posteriores de desarrollo. Una vez que un folículo primario deja la zona de almacenamiento deberá o seguir su desarrollo hasta la ovulación, o inducido a la atresia.

Figura 49. Estructura de un folículo primario.
El folículo primario es más grande que el folículo
primordial, la capa de células granulosas debajo de la membrana basal se hace
más prominente y aparece una estructura que acomapañará al óvulo por todo su
viaje llamada zona pellucida. En el ovocito puede observarse su bien formado
núcleo. Los óvulos liberados no se liberan de acompañantes después de ovular,
pues una capa de células granulosas se especializa para regular el desarrollo
del ovocito. Adicionalmente se desarrolla una capa semitransparente que media
la comunicación entre las células granulosas y el ovocito en desarrollo. Esta
membrana es conocida como zona pellucida. La estructura completa se denomina
primer estado del desarrollo folicular. En las siguientes dos etapas del
desarrollo folicular, las células granulosas se desarrollan y especializan.
El folículo secundario
El folículo en la etapa secundaria de su desarrollo ya está solo, su crecimiento se debe principalmente al crecimiento de las células de la capa granulosa, formando de este modo varias capas. A medida que el folículo va creciendo, las células granulosas próximas a la membrana basal se diferencian en dos capas denominadas teca, una la capa de teca interna y la otra es la teca externa. Las células de la capa teca median con los vasos sanguíneos, linfáticos y nervios.

Figura 50. Estructura de un folículo
secundario.
El folículo terciario
Las células de la teca interna eventualmente se hacen
planas, conformando una especie de epitelio escamoso y se especializan en la
producción de hormonas esteroides. Las células granulosas adquieren receptores
para ser activadas por medio de la hormona folículoestimulante y empiezan a
producir pequeñas cantidades de estrógenos. Las células de la teca externa
permanecen como fibroblastos y proveen un soporte estructural al folículo en
desarrollo. El desarrollo más allá del folículo primario es dependiente de la
presencia de la activación de los receptores de gonadotrofinas, inicia con la
menarquia en la pubertad y termina con la menopausia al final de la edad
reproductiva.
A medida que el folículo sigue creciendo, las células de la teca externa se expanden dejando abiertas cavidades llenas de fluido llamados antro al interior de las células granulosas. Una vez se ha formado el antro, el folículo ha ingresado en la etapa terciaria de su desarrollo. La hormona critica responsable de la progresión preantral y antral de un folículo es precisamente la hormona folículo estimulante. La hormona folículoestimulante induce la mitosis de las células granulosas. A medida que el número de células granulosas se incrementa, la producción de estrógenos, la capacidad de ser estimuladas por la hormona folículoestimulante, el tamaño del folículo y el volumen de fluido folicular también se incrementan.

Figura 51. Estructura del
folículo terciario.
El folículo de Graaf
A medida que el antro se hace más grande, el ovocito es
empujado a la periferia del folículo, en este estado de desarrollo se lo
denomina folículo de Graaf o folículo ovárico. Tres compartimentos diferentes
de células granulosas son evidentes en un folículo de Graaf. Las células
granulosas que rodean al ovocito se denominan cumulus grabulosa. Las células que se alinean formando el marco del
antro se denominan granulosas antrales. Las células que se encuentran en
contacto con las células teca se denominan granulosas del muro.
Las granulosas del muro y las granulosas del antro son más activas en la producción de estrógenos que las granulosas del cúmulo ovárico. Adicionalmente a la producción de hormonas endocrinas, los folículos de Graaf poseen un microambiente único, pues el líquido del antro posee diferentes concentraciones de hormonas de la pituitaria, esteroides, péptidos y factores de crecimiento. El fluido del antro puede tener concentraciones de 100 a 1000 veces más altas de estas hormonas que la sangre. El destino del folículo de Graaf está marcado por un evento determinante en su vida y es la ovulación, proceso por el cual el óvulo es liberado del folículo y del ovario mismo. El destino del óvulo normalmente se encuentra en las trompas de Falopio. Sin embargo, el folículo descartado no se degrada inmediatamente, en su lugar sigue una progresión subsecuente que lo transforma en el cuerpo lúteo. El destino del cuerpo lúteo está enlazado al del óvulo, si este no es fecundado el cuerpo lúteo de degrada rápidamente, pero su se da la gestación la vida del cuerpo lúteo se puede prolongar hasta el tercer mes de embarazo.

Figura 52. Un folículo secundario a la izquierda y el
folículo de Graaf a la derecha.
La degeneración de un folículo al interior del ovario se
denomina atresia y es caracterizada por la destrucción del ovocito y las células
granulosas. La atresia es un proceso continuo y puede ocurrir en cualquier
etapa del desarrollo folicular. Durante la vida de una mujer aproximadamente
entre 400 y 500 folículos logran llegar a la ovulación. Esta puede parecer una
cifra impresionante, pero en comparación con los 1 o dos millones de folículos
primordiales con los que la mujer nace se hace evidente algo, la mayoría de los
folículos pasan por la atresia y solo unos pocos logran llegar a la ovulación. La causa de la atresia es probablemente la
carencia de gonadotropinas que soporten el crecimiento folicular. Por ejemplo,
al inicio del ciclo menstrual una gran cantidad de folículos son seleccionados
para el crecimiento a folículo secundario, pero solo uno llamado “Folículo
Dominante” logra ovular. Debido a que el folículo dominante posee una fuente de
sangre preferencial, adquiere una mayor cantidad de hormona folículoestimulante
y hormona luteinizante. Otras razones para la atresia es que los folículos no
expresan suficientes receptores para las gonadotropinas o la incapacidad de las
células teca y/o granulosas de llevar a cabo la síntesis de esteroides hasta el
estradiol. Durante el proceso de la atresia, las células granulosas son
inducidas a la apoptosis, y/o el ovocito es inducido a una pseudomaduración.
Durante las primeras etapas de la pseudomaduración, la membrana nuclear se
desintegra, la cromatina se condensa y los cromosomas se agrupan como si fueran
a ir a la metafase. El término pseudomaduración se designa debido a que estos
ovocitos no son capaces de ser fertilizados exitosamente aun cuando puedan ser
ovulados. Durante la atresia de los folículos las células teca llevan a cabo
una hiperplasia e hipertrofia y pueden permanecer como una inflamación
folicular por un periodo de tiempo prolongado.
No hay comentarios:
Publicar un comentario