(Ciencias de Joseleg)(Biología)(Reproducción en los seres vivos)(Reproducción humana)(Introducción)(Historia de vida humana)(Introducción al sistema reproductor)(Regulación hormonal masculina)(Fisiología del sistema reproductor masculino)(Testosterona, masculinidad y reproducción)(La espermatogénesis humana)(Regulación hormonal femenina)(Anatomía del sistema reproductor femenino)(Los estrógenos, la feminidad y la reproducción)(La ovogénesis)(La ovulación y el cuerpo lúteo)(El ciclo menstrual)(El coito efectivo y el viaje de los espermatozoides)(De la fecundación a la implantación)(Gastrulación y formación de los discos embrionarios)(Los sacos embrionarios)(Destinos del disco trilaminar)(Gemelos y las membranas fetales)(Desarrollo fetal y embarazo)(Referencias bibliográficas)(Versión documento word)
En la sección anterior hemos visto como se forman las tres capas germinativas, así como otras estructuras accesorias como el canal neural y el notocordio. Ahora discutiremos el destino de cada uno de los discos germinativos. Hay que tener en cuenta que estos discos no siempre serán planos, y posteriormente el embrión se entuba.

Figura 85. Neuralización. Este evento es de vital importancia debido a sus implicaciones
éticas y morales sobre la posibilidad de percepción de dolor por parte del
embrión.
Al formar el tubo
el ectodermo rodeará a las otras dos capas, mientras que el endodermo formará a
su vez un tubo interno conectando dos orificios de salida. El mesodermo se
queda en medio para formar diferentes órganos. Una vez que los tres linajes
celulares se han formado dará inicio al periodo de organogénesis entre las
semanas 3 y 8 de embarazo. Cabe anotar que la mayoría de los embarazos son
diagnosticados en este periodo de tiempo debido a la exigencia cada vez mayor
del embrión por nutrientes de la madre y al cambio hormonal que esta sufre en
estos momentos.
Para el final de la
etapa de organogénesis los principales sistemas de órganos estarán
establecidos, así como la silueta de un cefalocordado reconocible de forma muy
clara. Al inicio de la tercera semana de desarrollo el epiblasto que de aquí en
adelante será denominado ectodermo posee la forma de un disco que es más ancho
en la región cefálica que en la región caudal. a aparición del mesodermo de la
notocorda y precordal inducen al mesodermo a endurecerse formando el plato
neural “recordemos de la notocorda se forma a partir del endodermo”. El plato
neural posee las células que se diferenciarán para formar la columna, iniciando
el proceso de neuralización.
Para el tiempo en
que el tubo neural se cierra, los tejidos oculares empiezan su desarrollo. En
términos generales la línea germinativa del ectodermo da lugar a órganos y
estructuras que mantienen el contacto con el ambiente externo. El sistema
nervioso central que mediante el mecanismo de neuralización se invagina al
interior del mesodermo. El sistema nervioso periférico gracias a la migración
de células de la cresta neural. El epitelio sensorial de las orejas, la nariz,
los ojos, el cuero cabelludo entre otros. Adicionalmente da lugar a las glándulas
subcutáneas, mamarias, pituitarias y al enamel de los dientes.
La neuralización es
el proceso por el cual se forma el tubo neural” a partir del plato neural. Para
el fin de la tercera semana las puntas laterales del plato neural se elevan
para formar los pliegues neurales y la región intermedia se hunde para formar
un canal que se cierra paulatinamente hasta formar un tubo. El tubo neural se
cierra aproximadamente en el día 25 junto con el desarrollo del tejido neural
en su interior, la medula espinal a lo largo del dorso y una zona más gruesa en
la región cefálica siendo esta el primordio cerebral.
Las células de la
cresta neural se encuentran en los labios del tubo neural antes de que este se
cierre. Esta población de células es posteriormente cubierta por el ectodermo,
y en este punto experimentan una transición muy fuerte, proliferando y dando
origen a otros grupos de tejidos: ganglios sensoriales, neuronas simpáticas y
entéricas, células de Schwann, células de la medula adrenal, también
contribuyen a la formación del esqueleto cráneo-facial, las neuronas de los
ganglios craneales, células gliales, melanocitos entre otros.
Las células de la cresta neural son tan fundamentales y tan versátiles en los tejidos a los cuales dan origen que algunos autores han propuesto denominarlas como una carta capa germinativa embrionaria. Evolutivamente estas células debieron aparecer cerca del origen de los vertebrados, es decir en el ancestro común de anfioxus y los demás vertebradoras.

Figura 86. Neuralización 2. La neuralización también hace que la organización del cuerpo
empiece a cambiar de un disco plano a una estructura similar a un gusano
segmentado, el proceso es semejante a la fabricación de una empanada.
Durante el
desarrollo de la gástrula el mesodermo no solo se almacena en las capas del
disco germinal, sino que se distribuye a lo largo de todo el embrión, tanto
hacia abajo cubriendo al endodermo y el saco vitelino como hacia arriba
cubriendo a las células del amnios. Aun así, la mayor parte del mesodermo se
almacena entre el ectodermo y el endodermo en el disco germinativo, esta masa
de mesodermo se denomina mesodermo paraxial “paralelo al eje anteroposterior”.
El mesodermo que cubre las células del amnios es denominado mesodermo parietal,
mientras que el que cubre al endodermo se denomina mesodermo visceral. Ambos se
agrupan como mesodermo somático. Adicionalmente una capa de mesodermo une al
paraxial y al somátido y se denomina mesodermo intermedio. Adicionalmente el
embrión como un todo adquiere una forma de 8, donde la zona más delgada
adquiere el nombre de cavidad intraembrionaria corporal “día 21”.
El mesodermo paraxial se caracteriza por su segmentación muscoloesquelética. Cada segmento se denomina somitomero o somita, sin embargo, con el ánimo de no introducir terminología extra nos referiremos a ellas como segmentos. Los segmentos aparecen primero en la región cefálica en asociación a la segmentación del plato neural en neuromeros. Una vez que aparece el primer segmento en la región cefálica, nuevos segmentos se adicionan a una tasa de tres por día. La primera aparece aproximadamente en el día 20 y para el final de la quinta semana se han desarrollado entre 42 y 44 pares de segmentos.
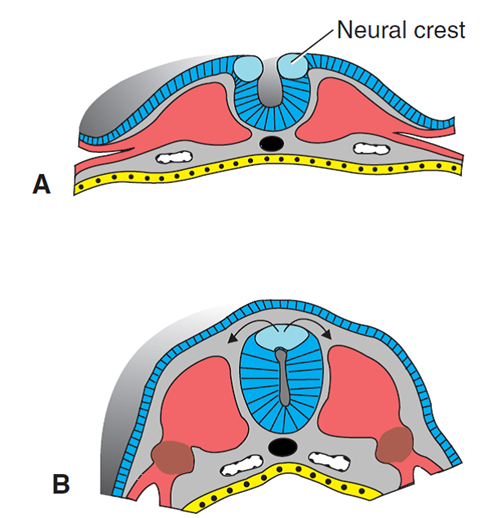
Figura 87. Neuralización 3. Cierre del tallo primitivo para formar la cresta
neural, el proceso deja una enorme cicatriz en el dorso.
Cada pareja se agrupa dependiendo de la región: 4 en la región occipital; 8 en la cervical; 12 en la torácica; 5 en la lumbar y entre 8 y 10 en la coxigeal. El primer segmento occipital y entre 5-7 de las coxigeales son retraídas, mientras que los segmentos restantes formaran el esqueleto del eje (axial). Debido a que los segmentos aparecen con una periodicidad predecible, es posible estimar la edad del embrión por la cantidad de segmentos que presenta.

Figura 88. Neuralización 4. En los modelos anteriores tenemos descripciones de: A día 17; B
día 19; C día 20; D día 21. En azul el ectodermo, en anaranjado el mesodermo y
en amarillo el ectodermo. La capa delgada de mesodermo en el disco germinativo
da lugar a una masa llamada mesodermo paraxial, del cual derivarán
posteriormente los segmentos corporales. Cabe anotar que la vista del embrión
en este punto es desde la cabeza a la cola.
Al inicio de la
cuarta semana de gestación las células del mesodermo de cada segmento empiezan
a experimentar especialización celular con una adecuada transformación de su
estructura y funciones. Algunas migran para rodear al tubo neural y la
notocorda, siendo denominados esclerotomo. Las células del esclerotomo
posteriormente se diferenciarán para formar las vértebras y los arcos de las
costillas. Las células del mesodermo de los segmentos ubicadas en los vértices
ventrolaterales y la parte superior son los precursores del sistema muscular
“miotomo” en su gran mayoría, así como la dermis dermatomo. Cada segmento actúa
independientemente, algo semejante a la organización de los gusanos
segmentados, de esta forma cada segmento desarrolla su propio esclerotomo, su
propio miotomo y su propio dermatomo. Adicionalmente cada segmento posee su
propia región neural en desarrollo.
El mesodermo intermedio conecta de manera temporal al mesodermo paraxial y al mesodermo lateral. El mesodermo intermedio se diferencia posteriormente en el sistema urogenital. La parte que será distribuida la parte torácica superior desarrolla grupos celulares que en el futuro serán nefrotomos. Mientras que los que se distribuyen de forma caudal “espalda” forman una masa de tejido no segmentada denominada cuerda nefrogénica. Las unidades del sistema excretor “riñones” y las gónadas se desarrollan en el mesodermo intermedio no segmentado.

Figura 89. Neuralización 5. Desde el exterior "ectodermo" la estructura del embrión
empieza a asemejarse a la de un gusano, también empiezan a aparecer los
segmentos.
El mesodermo del
plato lateral o somático se distribuye alrededor del amnios y del saco
vitelino, entrando en contacto con el endodermo y el mesodermo. Este mesodermo
se divide en dos, el parietal (somático) y el visceral (esplénico). Esta
diferenciación obedece precisamente con las cavidades y los tipos de tejido con
los que está en contacto. El mesodermo parietal emerge de aquel mesodermo
lateral que está en contacto con el ectodermo y el saco vitelino; mientras que
el mesodermo visceral emerge de aquel mesodermo del plato lateral que estaba en
contacto con el endodermo y la cavidad del amnios.
El mesodermo parietal formará las paredes laterales del cuerpo, así como sus pliegues. Estos pliegues junto con la cabeza y la cola se cierran en la pared corporal ventral. La capa parietal del plato lateral del mesodermo forma la dermis de la piel, las extremidades, los huesos, el tejido conectivo y el esternón. Adicionalmente, poblaciones celulares del ectodermo migran hacia el mesodermo, estas son el esclerotomo (formador de esqueleto) y el miotomo (formador de musculo) para formar los cartílagos de los costados, los músculos de las extremidades, y la mayor parte de los músculos superficiales del cuerpo.

Figura 90. Las somitas. La formación de los segmentos es paralelo al cierre
del tubo neural. Los segmentos de mesodermo se marcan claramente en el
ectodermo como si fueran musculo, lo que hace su identificación relativamente
fácil. La segmentalización ha sido uno
de los procesos más estudiados y en los cuales los genes Hox tienen una
importancia fundamental. Las señales para la diferenciación de los tejidos
emergen en cascada desde la notocorda, el tubo neural, la epidermis y el
mesodermo somático. Familias de genes Hox muy famosos se involucran en este
proceso como las noggin, los sonic hedgehog SHH y los genes PAX entre muchos
otros.
La capa visceral
del mesodermo del plato lateral junto con en endodermo embrionario forman las
paredes de tracto gastrointestinal. Las células del mesodermo de la capa
parietal rodean la cavidad intraembrionaria formando membranas muy delgadas: la
membrana mesotelial o serosa, las cuales enmarcan las futuras cavidades
celómicas “peritoneo, pleura y pericardio”. Se la denomina cerosa debido a que
segregan fluidos de naturaleza cerosa. Las células del mesodermo de la capa
visceral forman una delgada capa alrededor de cada órgano interno.
Las células de la
sangre y los vasos sanguíneos emergen del mesodermo. Los vasos sanguíneos se
desarrollan embrionariamente por dos mecanismos, la vasculogénesis y la
angiogénesis. En la vasculogénesis los vasos sanguíneos emergen de islas de
sangre, mientras que en la angiogénesis los nuevos vasos sanguíneos crecen a
partir de otros ya existentes.
La formación de la
sangre por vasculogénesis es representada por el modelo anterior, en este caso
se da por especialización celular de algunas céluulas del mesodermo llamadas
hemangioblastos que formaran las paredes de los primeros vasos sanguíneos, una
vez estos primeros se han formado el proceso de extensión del sistema
circulatorio se da por angiogénesis. Las primeras islas de sangre emergen en el
mesodermo que rodea al saco vitelino aproximadamente en la tercera semana de
gestación, y en pocas cantidades en el plato del mesodermo lateral y otras
regiones por vasculogénesis. Las islas emergen de las células del mesodermo
inducidas a especializarse en hemangioblastos, un precursor común de los vasos
sanguíneos y de la formación de la sangre. Aunque los primeros vasos sanguíneos
emergen de las islas de sangre en la pared del saco vitelino, esta población es
transitoria.
El tracto gastrointestinal es el órgano principal derivado del endodermo. Esta capa embrionaria cubre la superficie ventral del embrión y forma el techo del saco vitelino. Con el desarrollo de las vesículas cerebrales, el disco embrionario comienza inflamarse en la cavidad amniótica. Con el alargamiento del tubo neural el embrión se curva tomando la posición y la apariencia fetal vermiforme con unas regiones muy claras de cabeza y cola.


Figura 91. El sistema circulatorio. Las células madre hematopoyéticas
definitivas se derivan del mesodermo que rodea la aorta en un lugar cercano al
desarrollo del riñón mesofrénico llamada región aortagonadmesofrénica. Estas
células colonizan el hígado y se convierten en la mayor fuente hematopoyética
del embrión y el feto desde el segundo hasta el séptimo mes de gestación. Las
células madre del hígado colonizan luego el tuétano del hueso, que constituirá
el principal tejido hematopoyético desde el séptimo mes hasta la vejez.
Hasta este punto tenemos una empanada doble que se ha cerrado por el dorso, pero, aunque aún falta cerrar por el vientre, que es además la cavidad general más importante. El embrión en desarrollo con forma de 8 empieza alterar su estructura, la parte superior que está compuesta por la cavidad amniótica se extiende generando dos grandes pliegues llamados cavidad del cuerpo intraembrionario que rodean al cuerpo y lo encierran. El cierre se da en la posición ventral cortando al saco vitelino y dejando al embrión con dos cavidades generales.

Figura 92. Como leer los modelos. Vistas del embrión, los modelos de los embriones alternan entre
dos tipos de vista, a la izquierda tenemos una vista lateral del embrión como
si este fuera cortado desde la punta posterior hasta la anterior (desde la
cabeza a la cola) lo cual permite ver el interior de un lado del embrión, es
fácil reconocer esta vista ya que es la que permite ver el tallo conector que
posteriormente se convertirá en cordón umbilical. A la derecha tenemos una
imagen frontal del embrión como si alguien le hubiera cortado la cabeza y se pudiera
ver la parte media, esta vista puede reconocerse por el tubo neural en el
centro.
La primera cavidad
es formada por las cavidades embrionarias que rodean al sistema digestivo ya
encerrado, y la segunda es la cavidad amniótica que ahora rodea a todo el
embrión. En otras palabras, la cavidad amniótica ahora rodea al embrión. El
saco vitelino es cortado de la mayor parte del tracto gastrointestinal excepto
por la zona del tallo conector que de ahora en adelante será el cordón
umbilical. Falta mencionar la existencia de otro saco embrionario más pequeño
llamado alantoides, el cual también sufre el corte durante el cierre del
embrión, al igual que el saco vitelino el alantoides queda reducido a un ducto
al interior del cordón umbilical.
Como resultado del
crecimiento cefalocaudal y el cierre de la pared ventral del cuerpo, el
endodermo es incorporado al interior del cuerpo “al fin cobrando significado su
nombre como capa interna” formando el tracto gastrointestinal. El tracto
gastrointestinal se divide en tres zonas, superior, intermedio e inferior. El
tracto gastrointestinal intermedio se comunica con el saco vitelino “ahora
reducido a un tubo en el interior del cordón umbilical”. El tubo es amplio al
principio, pero se hace cada vez más del gado y largo a medido a que el embrión
crece y se desarrolla en el feto.
En la región
superior o cefálica el canal gastrointestinal se encuentra temporalmente unido
por una membrana denominada membrana orofaringeal. Esta membrana separa al
estomodeo de la faringe. El estomodeo es una estructura que se deriva del
ectodermo y terminará su desarrollo como la cavidad bucal; mientras que la
faringe es la punta del endodermo. Al ser estructuras que emergen de líneas
celulares diferentes, su acoplamiento puede tener problemas. El acoplamiento de
la cavidad bucal y de la faringe se da relativamente temprano durante el
embarazo, pues la membrana orofaringeal se rompe en la cuarta semana de
gestación, estableciendo un contacto permanente entre la boca y la faringe como
si fueran un solo canal.
El canal gastrointestinal inferior también termina en un límite de líneas celulares, por un lado, se encuentra el canal gastrointestinal que dará como resultado al intestino grueso y al recto que se derivan del endodermo, y por otro lado está el proctodeo, la cual es la parte final del recto y tiene su origen en el ectodermo. Al igual que en la anterior existe una membrana denominada membrana cloacal que separa a los dos tipos de tejido hasta que están listos para acoplarse.

Figura 93. Cierre ventral del
embrión y formación de las vísceras. Los modelos
anteriores nos muestran los cambios del embrión durante el proceso de cierre
ventral del embrión en una vista frontal. Arriba los pliegues laterales se
extienden, abajo a la izquierda los pliegues presionan al saco vitelino, abajo
a la derecha se muestra una zona donde los pliegues han cortado el saco
vitelino dejando solo la región del tracto gastrointestinal. Cabe anotar que no
todo el saco vitelino se pierde. Cuando el vientre se cierra el embrión se
enrosca adquiriendo la postura fetal típica, de todas las membranas
embrionarias el amnios aun continúa cubriendo el dorso "espalda" del
embrión.
El acoplamiento se
da en la séptima semana de gestación, en comparación con la del canal
bucofaríngeo que se da en la cuarta semana. Esta diferencia de tiempo sigue la
línea de desarrollo establecida por el canal neural y de cefalización, donde es
la cabeza la que se forma primero y la cola después. Otro resultado importante
del crecimiento cefalocaudal y del pliegue lateral es la incorporación parcial
del alantoides al cuerpo del embrión, conectado directamente a la cloaca. Desde
la cloaca el alantoides sigue su camino como un ducto en el interior del cordón
umbilical.
Aunque el saco
vitelino es una de las estructuras más emblemáticas de los amniotes “todos los
animales terrestres exceptuando a los anfibios”, su fisiología “función” sigue
siendo poco clara. Algunos autores estiman que su principal rol es el de servir
como órgano alimentario en las primeras etapas del desarrollo antes de la
implantación del embrión, función que sería mucho más vital en los animales
ovíparos que no experimentan la implantación del embrión.
Otros autores han apuntado a que contribuye a la formación de los primeros glóbulos rojos de la sangre, aunque esta función es altamente transitoria. Una de sus funciones primordiales es la de proveer las células madre de la línea germinal que se desarrollan en su pared posterior y que posteriormente migrarán hacia el mesodermo donde se desarrollan las gónadas. Estas células madre germinativa dependiendo del sexo se desarrollarán en óvulos o espermatozoides en la pubertad.

Figura 94. Cierre del embrión. Una vez llegados a este punto los modelos
embrionarios pasan de una vista anteroposterior a una vista lateral, Esto es
debido a que en el momento en que se cierra ventralmente, el embrión adquiere
ya una fisonomía semejante a un feto además de mostrar como el saco vitelino es
expulsado fuera de la estructura embrionaria.
A pesar de que el endodermo da lugar a estructuras tan importantes como el tejido endotelial del canal gastrointestinal, y las regiones intraembrionarias del saco vitelino y del alantoides, existen otras estructuras que tienen su origen en la línea germinativa del endodermo. Un ejemplo claro es el sistema respiratorio, el sistema respiratorio (laringe, tráquea y pulmones) tienen su origen evolutivo y embrionario en el canal gastrointestinal, por lo que su tejido endotelial también parte del endodermo. El parénquima de glándulas tan importantes como la tiroides, la paratiroides, el hígado y el páncreas. Tejido endotelial de la vejiga urinaria y la uretra. Dado que evolutivamente el canal auditivo tiene su origen en la cavidad bucal, el endodermo también hace parte del endotelio del canal auditivo.
1

2

3

Figura 95. Formación del sistema digestivo. (1) Cuando el cordón umbilical termina de formarse tanto el saco vitelino como el alantoides terminan por convertirse en secciones conectadas al tracto gastrointestinal en formación. De hecho, la posición del saco vitelino equivale a la posición del ombligo. En el modelo anterior podemos apreciar una vista lateral del embrión seccionado a la mitad, en amarillo se encuentra la estructura procedente del endodermo y en negro la del ectodermo. La cavidad bucal embrionario "estomodeo" penetra en el embrión hasta conectarse al endodermo; (2) El rostro embrionario permite ver con claridad la etapa de “pez”, y si ya sé que no deberíamos decir atapas, pero por favor no hay necesidad de ser sutiles cuando tenemos arcos branquiales más que evidentes; (3) La estructura anal del ser humano en el embrión recuerda a las cloacas de los reptiles, pues es una sola cavidad para el canal gastrointestinal y para el canal urinario en formación, sin embargo, en A la salida se encuentra obstruida por una membrana o proctodeo "closing plate", debido a esto la salida se encuentra por la ruta del alantoides hacia el cordón umbilical. B, C y D el mesodermo crece separando las cavidades y se forman los canales urinarios propiamente dichos. E desaparece el proctodeo y aparece el orificio urinario y el orificio anal.
No hay comentarios:
Publicar un comentario